Posted: January 15, 2009
The intelligent design creationists are jubilant -- a paper has been published that shows that organisms were front-loaded with genes for future function! It describes "'latent' or 'preexistent' evolutionary potential" in our history, they say.One small problem. The paper says nothing of the kind. It does mention latent potential, but it means something entirely different from something that is 'front-loaded', which is a sneaky little elision on the part of the creationists. There isn't even the faintest whiff of a teleological proposal in the paper at all, which makes me wonder if they even read it, or if, as seems more likely, they're simply incapable of comprehending the scientific literature.
So let's take a look at what the paper is actually about, and you'll see that it in no way supports the self-serving cheering of the creationists.
First, though, a little background that will be familiar to many of you, especially if you've read Gould's Full House: The Spread of Excellence from Plato to Darwin(amzn/b&n/abe/pwll). The process we're looking at is the increase in size and complexity of life over time, a subject near and dear to the heart of creationists who see it as a predestined pattern driven by intent, but also of great interest to legitimate biologists, who have found that no, it is not purposeful at all, but a natural consequence of chance variation.
Here's an analogy to get the right model into your head. Imagine a busy bar that closes at 2am, and sends all the drunks out the door to walk home. Since scienceblogs was so unfair to our Australian readership last night, let's imagine it is an Australian bar, and a million brain-blitzed Australian drunks spill out the door and start walking determinedly down the street. There are a few properties at play here. One is that this street happens to be paralleled on the right by a wall, so the drunks can't stagger too far in that direction. The other is that on the left is a wide-open sheep pasture which provides no obstacle to their progress that way. Another is that they are all initially aimed straight down the street, but because they are drunk, they stagger every once in a while and veer off a few degrees to the left or the right, entirely by chance.
You're hovering overhead in a helicopter. What do you think you will see?
The mob will proceed down the street, but as it goes, it will spread out gradually to the left. The majority will stagger right and left with equal frequency, and wobble roughly down the street. There will be a subset that will, by chance, stagger left a little more than to the right, and they'll drift off into the sheep pasture. Some may veer more to the right than the left, but they'll just bounce into the wall and get straightened out that way.
No drunk Australian has a preference to stroll into the sheep pasture. There is no intent to end up there. But some do, just by the odds. You, in your helicopter, can even look at the shape of the sprawling mob and make useful calculations about drunk Australian kinetics and make predictions about the aggregate trajectories of strolling drunkards, although you wouldn't be able to predict the pattern of an individual drunk.
This is the general model for how size and complexity vary over time. The direction of the street is time, and wandering out farther and farther into the sheep pasture is like getting larger or more complex. There are other details that the analogy does not cover, however. There may be advantages to wandering to the left -- it gets you out of the crowded mob of stumbling drunks. There may be intrinsic factors that limit how far a lineage can drift to the left, as well. Insect respiratory systems, for instance, create a kind of internal wall that limits how large they can get. There may also be external barriers that can be discerned by looking at the shape of the expanding mob.
Hop back into the helicopter and look down. What if the left edge of the mob doesn't expand exactly as you mathematically expected? What if it stops at some other barrier not visible from your vantage point? Perhaps there is a population of venomous Australian crocoducks lurking out there, or roving herds of carnivorous wallabies, and although we can't see them from up here, we do see that the drunks don't expand beyond a certain point.
Now that is the point of this new paper by Payne and others. They have taken a big picture examination of the distribution of fossil sizes over Earth's history, and asked whether the range has varied smoothly over time, as you'd expect if the outer bound were simply diffusing to higher levels. And the answer is no, it is not, there are a couple of discrete jumps in the maximum size that imply limits to earlier expansion that were overcome at specific periods in history. There was a wall of some sort to the left of the staggering mass of life, and they speculate a bit about what it might have been.
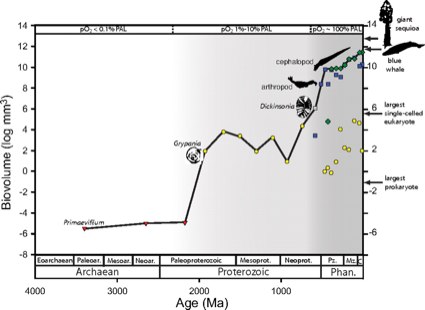
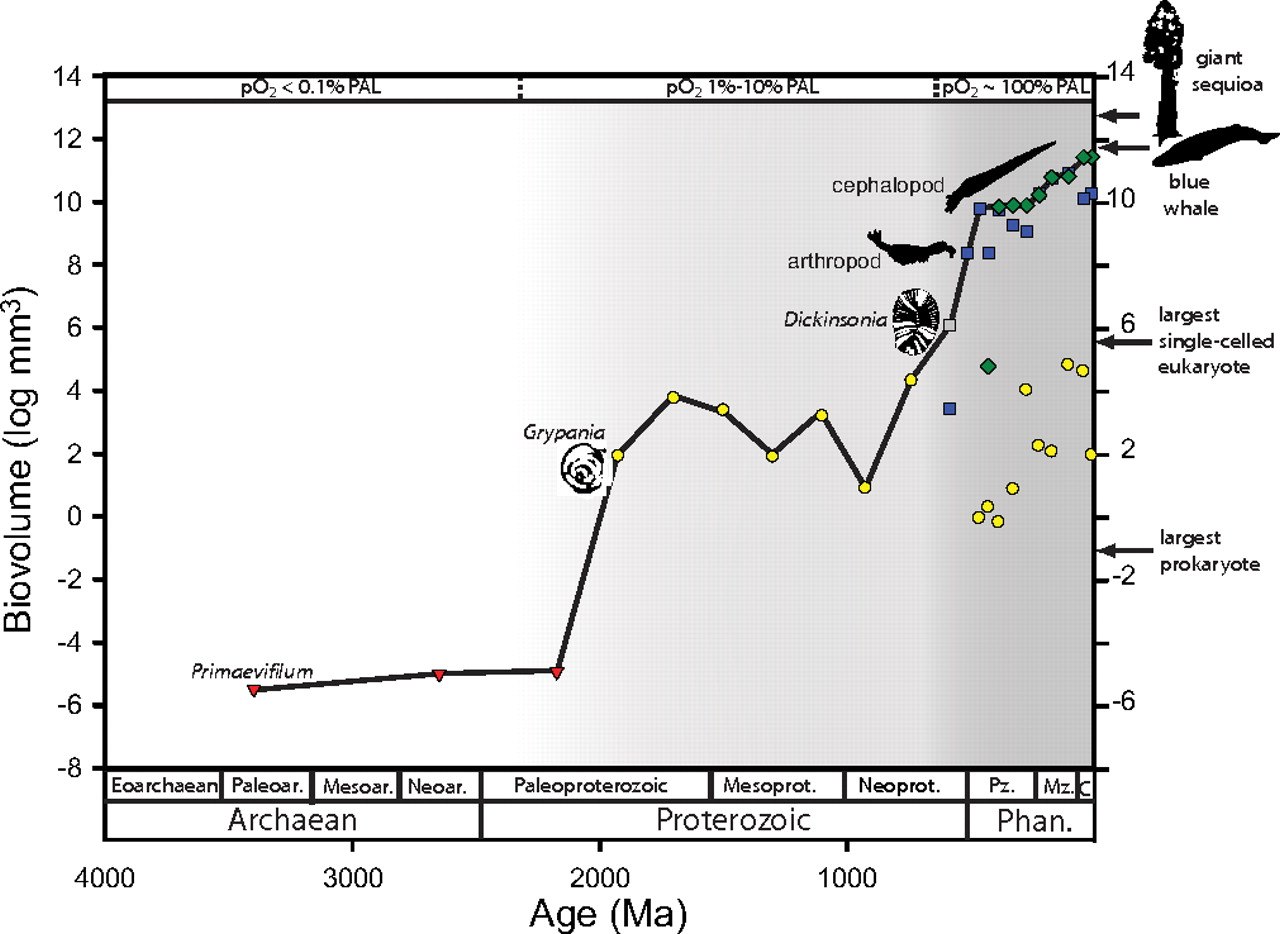
Here's the summary diagram of the results. The log of the volume of the largest fossils identified for a period are plotted on the Y axis, against time on the X axis. It is not a smooth curve, obviously -- there are two large upward lurches, one about 2 billion years ago and another over half a billion years ago.
Sizes of the largest fossils through Earth history. Size maxima are illustrated separately for single-celled eukaryotes, animals, and vascular plants for the Ediacaran and Phanerozoic. The solid line denotes the trend in the overall maximum for all of life. Increases in the overall maximum occurred in discrete steps approximately corresponding to increases in atmospheric oxygen levels in the mid-Paleoproterozoic and Ediacaran-Cambrian- early Ordovician. Sizes of the largest fossil prokaryotes were not compiled past 1.9 Gya. Estimates of oxygen levels from Canfield and Holland are expressed in percentage of PAL. Phan., Phanerozoic; Pz., Paleozoic; Mz., Mesozoic; C, Cenozoic. Red triangles, prokaryotes; yellow circles, protists; blue squares, animals; green diamonds, vascular plants; gray square, Vendobiont (probable multicellular eukaryote).How do we explain these sudden upward surges in maximum size? Unsurprisingly, it isn't by postulating a being of unimaginable magical or technological power who visits Earth at that time and inoculates his chosen species with size boosting genes. There is absolutely no evidence for that, and no need to invent such a silly hypothesis. Instead, there are two very good explanations that are actually supported by measurements and observations (and strangely, scientists prefer those kinds of explanations). One is a change in the environment, and the other is an intrinsic change in a subset of life.
The environmental change is illustrated in the chart. The periods when the maximum organism size increased are correlated with periods when free oxygen levels in the atmosphere increased. Basically, the atmosphere was modified by the byproducts of organic metabolism in a way that allowed aerobic organisms to grow to a larger size -- Earth accumulated enough rocket fuel in its atmosphere that some organisms could use to burn and grow. This is actually a fairly old story; we've been teaching about the oxygen increase in introductory biology for at least the last decade that I've been doing it.
The other part of the explanation is the one that has made our poor confused creationists so giddy, I'm afraid. The authors say,
These size steps coincide with, or slightly postdate, increases in the concentration of atmospheric oxygen, suggesting latent evolutionary potential was realized soon after environmental limitations were removed.
Ah, the realization of latent evolutionary potential. Did you know that you have latent evolutionary potential? Sure. If we put you and your family and friends in a novel environment, and let the generations tick by, we'll discover that certain sets of traits will become more prominent as selection and drift take their toll. The phrase does not imply that there is a purposeful arrangement of genes in your body that are there with a preexisting intent to allow you to thrive in a particular situation. You are complex, you have many properties that may in your current situation be superfluous or useless, but could be utilized in different situations.
You see, we have a good idea of exactly what intrinsic capabilities contributed to the 'latent potential' that led to certain lineages growing larger at those two transitions, and they both have natural precursors. We don't need a designer to explain the shifts, because the changes are expressions of known properties!
The abrupt increase in the Paleopterozoic, for instance, is the product of growth in size of the relatively recently evolved eukaryotes. You've probably heard of the endosymbiote hypothesis: eukaryotes are the product of a merger of multiple prokaryotic organisms into a single whole. Single celled organisms combined, with different specializations -- organelles in our cells called mitochondria, for instance, are thought to be descendants of an incorporated prokaryote. Our mitochondria have the primary function of burning carbon and oxygen to produce energy. This was "latent evolutionary potential" that could be exploited by eukaryotes as oxygen availability rose. And so, following the rise of increasing oxygen concentrations, the size of some eukaryotes staggered upwards to a new maximum.
The second surge in the beginning of the Phanerozoic was also a consequence of a property with precursors in the existing single-celled populations: multicellularity. This is another biological property rich in "latent evolutionary potential". Again and unsurprisingly, simply combining multiple cells into one organism is a fast-track to larger organismal size, and we see multiple lineages exploring this capability, many of which failed and died out, such as the Ediacaran fauna, and others in the Cambrian that expanded rapidly. Multicellularity itself is not an abrupt, binary choice. We have precursors: modern choanoflagellates show that protists can find selective advantage in transient assemblies, colonial organisms show the virtues of more permanent arrangements, and creatures like sponges exhibit cooperativity and specialization in internal function. Chance creates the potential, and selection can drive an agency-free promotion of greater expression of that potential.
I must emphasize that this is not a paper endorsing any form of intelligent design creationism, and the creationists' appropriation of its conclusion depends entirely on their distortion of its contents. Here is the authors' full conclusion.
Although increase in maximum size over time can often be accounted for by simple diffusive models, a single diffusive model does not appear capable of explaining the evolution of life's overall maximum size. Approximately 3/4 of the 16-orders-of-magnitude increase in maximum size occurred in 2 discrete episodes. The first size jump required the evolution of the eukaryotic cell, and the second required eukaryotic multicellularity. The size increases appear to have occurred when ambient oxygen concentrations reached sufficient concentrations for clades to realize preexisting evolutionary potential, highlighting the long-term dependence of macroevolutionary pattern on both biological potential and environmental opportunity.
The way the creationists have abused that is by pretending that this implies that the evolution of the eukaryotes and of multicellularity had to have been purposeful events. This is simply not true. All this is saying is that the limits of growth are properties of both organismal and environmental constraints, and that we can map those out by looking at the fossil record.
Payne JL, Boyer AG, Brown JH, Finnegan S, Kowalewski M, Krause RA Jr, Lyons SK, McClain CR, McShea DW, Novack-Gottshall PM, Smith FA, Stempien JA, Wang SC (2009) Two-phase increase in the maximum size of life over 3.5 billion years reflects biological innovation and environmental opportunity. Proc Natl Acad Sci U S A. 106(1):24-7.
{kind=link}